Ke Guo1,
Gang Yin2 ![]() ,
Xiong-Hong Zi1,
Wen-Guang Yan3
,
Xiong-Hong Zi1,
Wen-Guang Yan3
For correspondence:- Gang Yin Email: docgk79@sina.com
Received: 16 April 2015 Accepted: 28 December 2015 Published: 28 February 2016
Citation:
Guo K, Yin G, Zi X, Yan W.
A Study on ex
© 2016 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To investigate the ex
Methods: Ischemia-reperfusion model was established by thread-occluded method. Tetrazolium red (TTC), H/E and Nissl staining were used to evaluate whether ischemia-reperfusion model was successfully established. TUNEL staining and immunohistochemistry were employed to monitor apoptosis-positive nerve cells as well as STAT3-, p-Tyr705-STAT3-, Bcl-2- and Fas-positive cells in ischemic penumbra (IP) and ischemic core (IC).
Results: The results of TTC, HE and Nissl staining indicated that the ischemia-reperfusion model was successfully established. After 3 h, ischemia followed by different reperfusion times, the STAT3-, p-Tyr705-STAT3-, Fas- and Bcl-2-positive cells counts and the apoptosis-positive nerve cells count were significantly (p < 0.05 or 0.01) increased to 27.20, 29.20, 15.90, 18.50, and 202.00 in IP and 19.50, 21.20, 12.50, 12.40, and 97.80 in IC, compared with the sham-operated group. As reperfusion times increased, cell counts did not decrease significantly relative to control group. Correlation analysis indicate that there was significant (p < 0.01) positive correlations among STAT3-, p-Tyr705-STAT3-, Fas- and Bcl-2-positive cells counts on the one hand, and apoptosis-positive nerve cells count in IP and IC, on the other hand.
Conclusion: Regulating ex
Introduction
Cytokines and inflammatory mediators such as interleukin-6 (IL-6), interleukin-11 (IL-11), leukemia inhibitory factor (LIF) and ciliary neurotrophic factor (CNTF) are a few key factors in ischemic cerebrovascular disease [1-3]. Janus kinase-signal transducers and activators of transcription (JAK-STAT) signaling pathway plays an important role in regulating IL-6, IL-11, LIF, CNTF, proliferation, differentiation, apoptosis and immunoregulation [4-9]. It is reported that STAT3, a STAT family member, can mediate apoptosis by regulating the expression levels of Bcl-2 and Bcl-xl, but Fas-mediated cell apoptosis signal inhibits the expression and phosphorylation of STAT3 [10,11]. It is reported that the expressions of STAT3 were increased after focal cerebral ischemia [12], but the mechanism is still unknown. Additionally, although the effect of focal cerebral ischemia on the STAT3 phosphorylation in ischemic penumbra (IP) and ischemic core (IC) has been reported [13], the effect of STAT3 phosphorylation on cell apoptosis and the correlations among STAT3 phosphorylation, cell apoptosis and apoptotic proteins (Bcl-2 and Fas) in focal cerebral ischemia are still unknown.
In this study, the effects of the STAT3 and its phosphorylation on neuronal apoptosis in focal cerebral ischemia-reperfusion rat model were investigated by detecting the expression levels of STAT3, STAT3 tyrosine 705 phosphorylation (p-Tyr705-STAT3), Fas, Bcl-2 and the apoptosis-positive nerve cells count and analyzing the correlations among them.
Methods
Chemicals and reagents
Rabbit anti STAT3 and p-Tyr705-STAT3 polyclonal antibodies were obtained from Santa Cruz (Santa Cruz, USA). Rabbit anti Bcl-2 and Fas antibodies, Ready-to-use strept avidin-biotin complex (SABC) immunohistochemical kit, phosphate buffer solution (PBS) and polylysine were purchased from BOSTER (Wuhan, China). Diaminobenzidine (DAB) chromogenic reagent kit and TUNEL staining kit were obtained from Zhongshan Biotechnology (Beijing, China). Tetrazolium red (TTC) was purchased from Sigma (St. Louis, MO, USA).
Animals
Healthy, adult and male Wistar rats (300 ± 20 g) were provided by Laboratory Animal Centre, Xiangya School of Medicine, Central South University, Changsha, China. All procedures involving animals were conducted in accordance with the National Institute of Health Guide for the Care and Use of Laboratory Animals [14]. All experiments were performed with the approval of Ethics animals of Central South University in this study (protocol no. TXHCSU10491). Animals were housed in a room with the temperature of 25 ± 2 ºC, relative humidity of 50 - 60 % and 12 h light/dark cycle, and had free access to food and water.
Establishment of focal cerebral ischemia-reperfusion rat model
Focal right middle cerebral artery ischemia-reperfusion rat model was established by thread-occluded method [15,16]. Rats were firstly anaesthetized by 10 % chloral hydrate (0.35 mL/100 g) intraperitoneally at 37 ± 0.5 ºC and then fixed on surgery board in the supine position. A midline incision on the neck was made to expose the external carotid artery (ECA), right common carotid artery (RCCA) and internal carotid artery (ICA). Then the ECA and RCCA were ligated. The blood flow of ICA was blocked by pulling the preset string, and a V-notch between bifurcation and the ligation of RCCA was cut. The nylon thread (length: 5.5 cm; diameter: 0.25 - 0.28 mm) was carefully introduced into the ICA lumen through the V-notch until 2.0 - 2.3 cm. The ICA was ligated to prevent the bleeding for 3 h. Then bleeding was re-perfused for appropriate time by withdrawing the nylon thread. Sham-operated group were performed with the same surgical exposure procedures, but the carotid arteries were not blocked.
Animal grouping and treatment
45 Wistar rats were randomly divided into 9 groups (n = 5): normal group (NG), sham-operated group (SG) and seven focal cerebral ischemia-reperfusion model groups (MG + 0.5 h, 3 h, 6 h, 12 h, 24 h, 3 days or 7 days reperfusion). After observing, rats were anaesthetized by 10 % chloral hydrate (0.35 mL/100 g) intraperitoneally and sacrificed.
Neurological defect scoring
After operation, the neurological statuses of rats were scored on a five-point scale [17]. Rats with no neurologic impairment scored 0. Left forelimb of rats with adduction, flexion and failure to fully stretch scored 1. Rats with circling and rotating to left when crawling scored 2. Rats with toppling to the contralateral side when standing scored 3. Rats with failure to walk spontaneously scored 4.
TTC, H/E, Nissl and TUNEL staining
After rats were sacrificed, their brains were harvested and frozen at -20 ºC. After 20 min, the coronal plane was obtained from brain via a tissue slicer (AO, USA) and stained with 2 % TTC staining solution at 37 ºC for 30 min. The remaining brain tissue samples were fixed in 4 % paraformaldehyde for 6 h. Then the IP of the fixed brain tissue was treated with the following steps: dehydration, transparency, dip in wax, embedding and slicing. The paraffin sections were stained by Harris hematoxylin and eosin (H/E) staining solution, 1 g/L Nissl staining solution and TUNEL staining kit, respectively. Subsequently, the H/E staining, Nissl staining and TUNEL staining slicing were observed and photographed by microscope. The apoptosis-positive nerve cells count in TUNEL staining slicing was analyzed by HPIAS-100 high-definition color pathological image analysis system (Wuhan, China).
Immunohistochemical staining for STAT3, p-Tyr705-STAT3, Bcl-2 and Fas
The coronal plane was obtained from the fixed brain tissue, and then focal side and uninjured side were picked out from coronal plane. Further, IC and IP were picked out from focal side, and meanwhile, the counterpart of the ischemic brain (CIB) was obtained. According to method as above described, the different tissues were made into paraffin section. After heat-mediated antigen retrieval, paraffin sections were incubated with STAT3, p-Tyr705-STAT3, Bcl-2 and Fas antibodies and subsequently with SABC immunohistochemical kit. After staining by DAB chromogenic reagent kit, the paraffin sections were observed and analyzed by microscope and HPIAS-100 high-definition color pathological image analysis system (Wuhan, China).
Statistical analysis
All data are presented as mean ± standard deviation. Differences among different groups were analyzed by one-way ANOVA (LSD or Games-Howell test) on SPSS 21.0. Correlations among the apoptosis-positive nerve cells count and the STAT3-, p-Tyr705-STAT3-, Bcl-2- and Fas-positive cells counts in IP and IC were analyzed by two-tails Pearson or Spearman test on SPSS 21.0. When p-value was lower than 0.05 or 0.01, differences were considered statistically significant.
Results
Neuro-behavioral observation
Neurological defect scoring was used to evaluate whether the focal cerebral ischemia modeling was successfully established. When the neurological defect scoring was 1, 2 or 3, rat was selected out as experimental animals. When the neurological defect scoring was 0 or 4, it indicated that the focal cerebral ischemia modeling was unsuccessfully established. NG and SG did not have neurological impairment. The unqualified focal cerebral ischemia rats were replaced with standby animals.
TTC staining
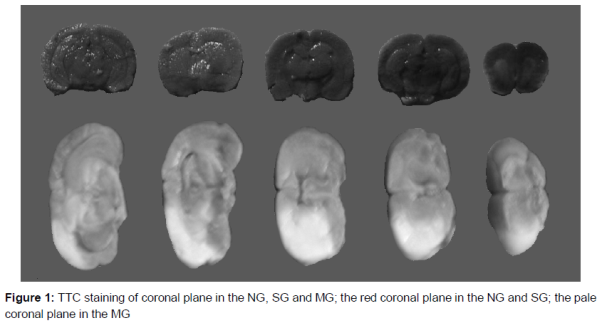
After staining with TTC, the color of ischemic tissue was pale, and the color of normal tissue was red. As shown in , the color of the coronal plane in the NG and SG was red, and the color of the coronal plane in the MG was pale.
H/E and Nissl staining
As shown in , the form and construction of brain nerve cells, spongiocyte and capillaries in NG and SG were normal and intact, and there was no red dye in the cytoplasm. After 3 h ischemia followed by 0.5 or 3 h reperfusion, there was not evident changes in the form and construction of brain nerve cells, spongiocyte and capillaries. With the increase of reperfusion time, necrocytosis and cell losses were evidently observed. When the reperfusion time was 12 h, the mild interstitial edema was observed. When the reperfusion time was 24 h, the moderate interstitial edema, the reduction of cell count, the swelling and irregular cellular morphology and the inflammatory cell infiltration were evidently observed in HE staining. Furthermore, the karyopyknosis, the shrink and deformation of soma, the losses of Nissl’s body and ghost cell were evidently observed in Nissl staining. When the reperfusion time was 3 or 7 days, extensive neuronal deaths near the pyramidal cells and corpus striatum of cerebral cortex were evidently observed.
TUNEL staining
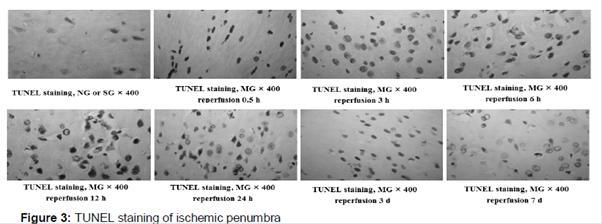
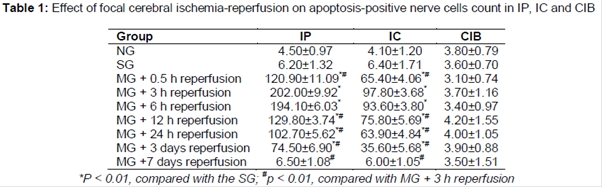
As shown in and , the apoptosis-positive nerve cells counts were not significantly different between the NG and SG, and it indicated that the apoptosis-positive nerve cells count was not affected by the sham-operation. After 3 h ischemia followed by different reperfusion times, the apoptosis-positive nerve cells counts in IP and IC were significantly (p < 0.01) increased and then decreased to no significant difference, compared with the SG. Additionally, there were a few apoptosis-positive nerve cells were detected in CIB of the NG, SG and MG.
Effect of focal cerebral ischemia-reperfusion on STAT3-, p-Tyr705-STAT3-, Bcl-2- and Fas-positive cells counts
As shown in Tables 2 - 5, the STAT3-, p-Tyr705-STAT3-, Fas- and Bcl-2-positive cells counts were not significantly different between the NG and SG, and it indicated that the STAT3-, p-Tyr705-STAT3-, Fas- and Bcl-2-positive cells counts were not affected by the sham-operation. After 3 h ischemia followed by different reperfusion times, the STAT3-, p-Tyr705-STAT3-, Fas- and Bcl-2-positive cells counts in IP and IC were significantly (p < 0.05 or 0.01) increased and then decreased to no significant difference, compared with the SG. Additionally, there were a few STAT3-, p-Tyr705-STAT3- and Bcl-2-positive cells were detected in CIB.
Correlations among the STAT3-, p-Tyr705-STAT3-, Bcl-2- and Fas-positive cells counts and the apoptosis-positive nerve cells count in IP and IC
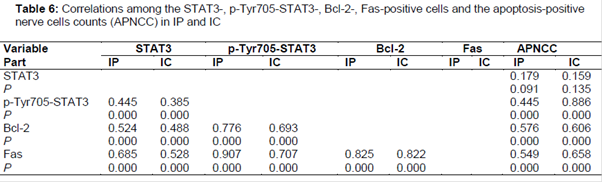
The correlations among STAT3-, p-Tyr705-STAT3-, Bcl-2- and Fas-positive cells counts and the apoptosis-positive nerve cells count in seven focal cerebral ischemia-reperfusion model groups (MG + 0.5 h, 3 h, 6 h, 12 h, 24 h, 3 days or 7 days reperfusion) were analyzed by two-tails Pearson or Spearman test. As shown in , there are significantly (p < 0.01) positive correlations among STAT3-, p-Tyr705-STAT3-, Fas- and Bcl-2-positive cells counts in IP and IC, and the p-Tyr705-STAT3-, Fas- and Bcl-2-positive cells counts in IP and IC were significantly (p < 0.01) and positively related to the apoptosis-positive nerve cells count.
Discussion
The focal cerebral artery ischemia-reperfusion rat model is a commonly used model to investigate ischemia-reperfusion-related problems [18]. TTC, HE and Nissl staining are three commonly used methods to explore whether the focal middle cerebral artery ischemia-reperfusion rat model is successfully established [19,20]. The results of TTC, HE and Nissl staining indicated that the focal middle cerebral artery ischemia-reperfusion rat model was successfully established.
Ischemia brain tissue can be divided into IP and IC, whose sizes are in dynamic change [21]. Generally, necrosis is the main pathway of nerve cell in IC, but apoptosis is the main pathway of nerve cell in IP [22,23]. The necrosis and apoptosis in cerebral ischemia are related to the duration or degree of ischemia [17,24]. In the present study, TUNEL staining was used to study the effect of ischemia-reperfusion on the apoptosis-positive nerve cells count in IP and IC. The results of TUNEL staining indicated that after focal cerebral artery ischemia, the apoptosis-positive nerve cells count was increased in IP and IC, and the increase was reversed by reperfusion.
The inhibition of JAK-STAT signaling induces the apoptosis of cancer cell apoptosis via death receptor- and mitochondria-mediated apoptotic pathways [25,26]. Fas, a pro-apoptotic protein, plays an important role in death receptor-mediated apoptotic pathway [27]. Bcl-2, an anti-apoptotic protein, plays an important role in mitochondria-mediated apoptotic pathway [28]. The increase of the expressions of STAT3 and p-Tyr705-STAT3 proteins can induce the JAK-STAT signaling. In the present study, after 3 h ischemia followed by different reperfusion times, the expressions of STAT3, p-Tyr705-STAT3, Fas and Bcl-2 proteins were first increased and then decreased in IP and IC. The correlation analysis indicated that there are positive correlations among STAT3, p-Tyr705-STAT3, Fas and Bcl-2-positive cells counts, and the p-Tyr705-STAT3, Fas and Bcl-2-positive cells counts were positively related to the apoptosis-positive nerve cells count. These results indicated that the reversed mechanisms of reperfusion on the ischemia-induced nerve cells apoptosis were related to the interactions among STAT3, p-Tyr705-STAT3, Fas and Bcl-2 proteins.
Conclusion
The findings of this study show that regulating expression and tyrosine 705 phosphorylation of STAT3 may be a new effective strategy for treating cerebral infarction. Furthermore, the findings to further investigate the molecular pathological mechanism of focal cerebral ischemia.
References
Archives


News Updates